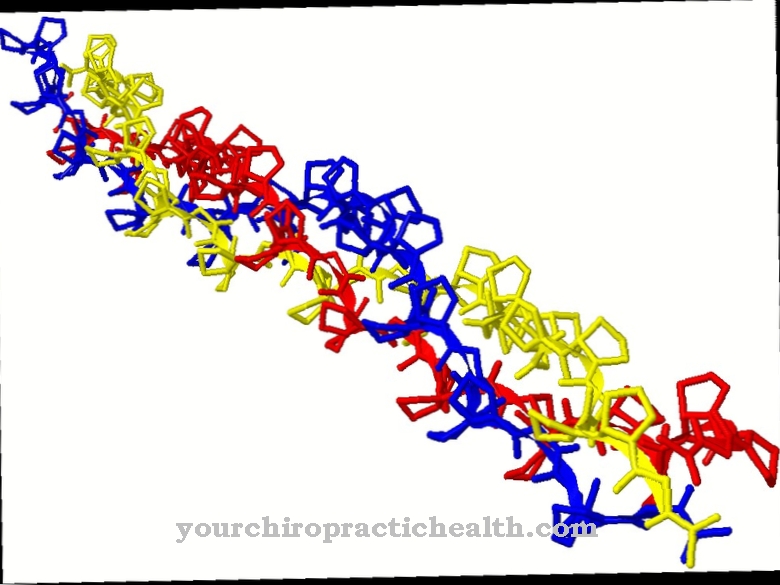
คู่เบสประกอบด้วยนิวคลีโอเบสสองตัวซึ่งตั้งอยู่ตรงข้ามกันในกรดดีออกซีไรโบนิวคลีอิก (DNA) หรือกรดไรโบนิวคลีอิก (RNA) จับกันและสร้างเกลียวคู่ด้วยความช่วยเหลือของพันธะไฮโดรเจน นี่คือข้อมูลจีโนมของสิ่งมีชีวิตและมียีน ข้อผิดพลาด การจับคู่ฐาน สามารถนำไปสู่การกลายพันธุ์
การจับคู่ฐานคืออะไร?
คู่ฐานประกอบด้วยนิวคลีโอเบส มันเป็นองค์ประกอบของ DNA หรือ RNA ในทางกลับกันนิวคลีโอเบสเหล่านี้ร่วมกับกรดฟอสฟอริกหรือฟอสเฟตและดีออกซีไรโบสซึ่งเป็นน้ำตาลทำให้เกิดนิวคลีโอไทด์ (เบส)
กรดฟอสฟอริกและดีออกซีไรโบสเหมือนกันสำหรับทุกนิวคลีโอไทด์พวกมันเป็นกระดูกสันหลังของดีเอ็นเอ ฐานและ deoxyribose เรียกว่านิวคลีโอไซด์ กากฟอสเฟตหมายความว่าดีเอ็นเอมีประจุลบและยังชอบน้ำอีกด้วยมันทำปฏิกิริยากับน้ำ
นิวคลีโอไทด์แตกต่างกันเฉพาะในฐาน ความแตกต่างเกิดขึ้นระหว่างห้าฐานขึ้นอยู่กับว่าเป็นส่วนประกอบของ DNA หรือ RNA ฐานคืออะดีนีน (A) และกัวนีน (G) ซึ่งเป็นของพิวรีน ไทมีน (T), ไซโตซีน (C) และยูราซิล (U) เป็นไพริมิดีน พิวรีนเป็นสารประกอบอินทรีย์เฮเทอโรไซคลิกในขณะที่ไพริมิดีนเป็นเฮเทอโรไซคลิกอะโรมาติกสารประกอบอินทรีย์
ในดีเอ็นเอมีการจับคู่เบสของอะดีนีนและไทมีน (A-T) เช่นเดียวกับกัวนีนและไซโตซีน (G-C) ในกรณีของ RNA ในทางกลับกันการจับคู่พื้นฐานเกิดขึ้นระหว่าง adenine และ uracil (A-U) และระหว่าง guanine และ cytosine (G-C) การจับคู่ฐานนี้เรียกว่าการเสริมกัน
การจับคู่ถูกสร้างขึ้นโดยพันธะไฮโดรเจน นี่คือปฏิสัมพันธ์ระหว่างอะตอมของไฮโดรเจนกับอิเล็กตรอนคู่เดียวบนอะตอมอื่น อะตอมของไฮโดรเจนมีพันธะโควาเลนต์ที่นี่ นี่คือพันธะเคมีที่มีปฏิสัมพันธ์ระหว่างเวเลนซ์อิเล็กตรอนของอะตอมหนึ่งกับนิวเคลียสของอะตอมอื่น คู่เบสยังใช้เป็นตัววัดขนาดของดีเอ็นเอ: 1bp สอดคล้องกับหนึ่งและ 1kb สอดคล้องกับ 1,000 คู่เบสหรือนิวคลีโอไทด์
ฟังก์ชันและงาน
การจับคู่ฐานมีหน้าที่ที่จำเป็นสำหรับโครงสร้างของดีเอ็นเอ ดีเอ็นเอเกิดขึ้นเป็นเกลียวคู่ การจัดเรียงเชิงพื้นที่ของเกลียวคู่เรียกว่า B-DNA ซึ่งเป็นเกลียวสองเกลียวทางขวาซึ่งตรงกันข้ามกับรูปตัว A มีการจัดวางที่ผ่อนคลายกว่า
เมื่ออะดีนีนและไทมีนจับคู่กันจะเกิดพันธะไฮโดรเจนสองพันธะ ในทางตรงกันข้ามการจับคู่กัวนีนและไซโตซีนจะสร้างพันธะไฮโดรเจนสามพันธะ เนื่องจากการจับคู่เบสระหว่างพิวรีนและไพริมิดีนระยะห่างระหว่างสายดีเอ็นเอทั้งสองจึงเท่ากันเสมอ ดีเอ็นเอมีโครงสร้างปกติเส้นผ่านศูนย์กลางของเกลียวดีเอ็นเอคือ 2 นาโนเมตร การหมุนที่สมบูรณ์ 360 °ภายในเกลียวเกิดขึ้นทุกๆ 10 คู่เบสและยาว 3.4 นาโนเมตร
การจับคู่ฐานยังมีบทบาทสำคัญในการจำลองแบบดีเอ็นเอ การจำลองแบบดีเอ็นเอแบ่งออกเป็นระยะเริ่มต้นระยะการยืดตัวและระยะสิ้นสุด สิ่งนี้เกิดขึ้นระหว่างการแบ่งเซลล์ DNA ถูกคลายออกโดยเอนไซม์ที่เรียกว่า DNA helicase เส้นคู่ถูกแยกออกจากกันและ DNA polymerase จะยึดตัวเองกับสาย DNA เส้นเดียวและเริ่มสร้างสาย DNA เสริมในแต่ละเส้นสิ่งนี้ทำให้เกิดดีเอ็นเอสายเดี่ยวใหม่สองเส้นซึ่งรวมกันเป็นเกลียวคู่ดีเอ็นเอใหม่
โครงสร้างของเกลียวคู่ดีเอ็นเอที่สังเคราะห์ขึ้นใหม่ได้รับการรับรองโดยการจับคู่เบสเสริม นอกจากนี้การจับคู่เบสยังมีบทบาทสำคัญในการสังเคราะห์โปรตีน ซึ่งแบ่งออกเป็นการถอดความและการแปล ในระหว่างการถอดความเกลียวคู่ของดีเอ็นเอจะถูกคลายออกและเส้นเสริมจะถูกแยกออกจากกัน สิ่งนี้ทำได้โดยเอนไซม์เฮลิเคส
พอลิเมอเรส RNA จับกับดีเอ็นเอสายเดี่ยวและสร้าง RNA เสริมกับมัน ในกรณีของ RNA จะใช้ uracil แทน thymine และเมื่อเทียบกับ DNA แล้วจะมีสิ่งที่เรียกว่า polyA tail RNA จะลงท้ายด้วย adenine เสมอ RNA ยังคงเป็นเส้นใยเดี่ยวและใช้ในการสังเคราะห์โปรตีนระหว่างการแปล ประเภทของโปรตีนขึ้นอยู่กับยีนเฉพาะที่ถูกอ่านและใช้เป็นแม่แบบในการสังเคราะห์โปรตีน
คุณสามารถหายาของคุณได้ที่นี่
➔ยารักษากล้ามเนื้ออ่อนแรงความเจ็บป่วยและความเจ็บป่วย
Erwin Chargaff พบว่าจำนวนของเบสอะดีนีนและไทมีนรวมทั้งกัวนีนและไซโตซีนเท่ากับ 1: 1 James D.Watson และ Francis Harry Compton Crick ในที่สุดก็พบว่ามีการจับคู่ฐานเสริมของอะดีนีนและไทมีนรวมทั้งกัวนีนและไซโตซีน สิ่งนี้เรียกว่าการจับคู่วัตสัน - คริก
อย่างไรก็ตามความผิดปกติต่างๆอาจทำให้เกิดการจับคู่พื้นฐานที่ผิดปกติเช่นการจับคู่วัตสัน - คริกแบบย้อนกลับ อีกรูปแบบหนึ่งของการจับคู่พื้นฐานที่ผิดพลาดคือการจับคู่แบบโยกเยก สิ่งเหล่านี้เป็นการจับคู่ที่ตรงกันข้ามกับการจับคู่วัตสัน - คริกเช่น G-U, G-T หรือ A-C ข้อผิดพลาดเหล่านี้อาจเกิดขึ้นได้ในระหว่างการจำลองแบบดีเอ็นเอและจากนั้นจะต้องถูกกำจัดโดยการซ่อมแซมดีเอ็นเอ
การจับคู่ฐานที่ไม่ถูกต้องอาจนำไปสู่การกลายพันธุ์ การกลายพันธุ์เหล่านี้ไม่จำเป็นต้องเป็นอันตราย มีสิ่งที่เรียกว่ามิวเทชั่นแบบเงียบซึ่งมีการแลกเปลี่ยนคู่เบสเป็นคู่อื่น แต่สิ่งนี้ไม่ส่งผลให้เกิดการรบกวนการทำงานหรือโครงสร้างของโปรตีนสังเคราะห์ อย่างไรก็ตามในกรณีของโรคโลหิตจางชนิดเคียวการกลายพันธุ์เป็นสาเหตุของการสร้างเซลล์เม็ดเลือดแดงที่ไม่ทำงาน การกลายพันธุ์ส่งผลโดยตรงต่อฮีโมโกลบินซึ่งมีหน้าที่ในการขนส่งออกซิเจนในเลือด ความผิดปกติของระบบไหลเวียนโลหิตที่ร้ายแรงและเป็นอันตรายถึงชีวิตและโรคโลหิตจางเกิดขึ้น









.jpg)



.jpg)





.jpg)